Contributors
Peter Thompson1
Karlie McDonald1
1 CSIRO Oceans and Atmosphere, Hobart, TAS, Australia
Key Information
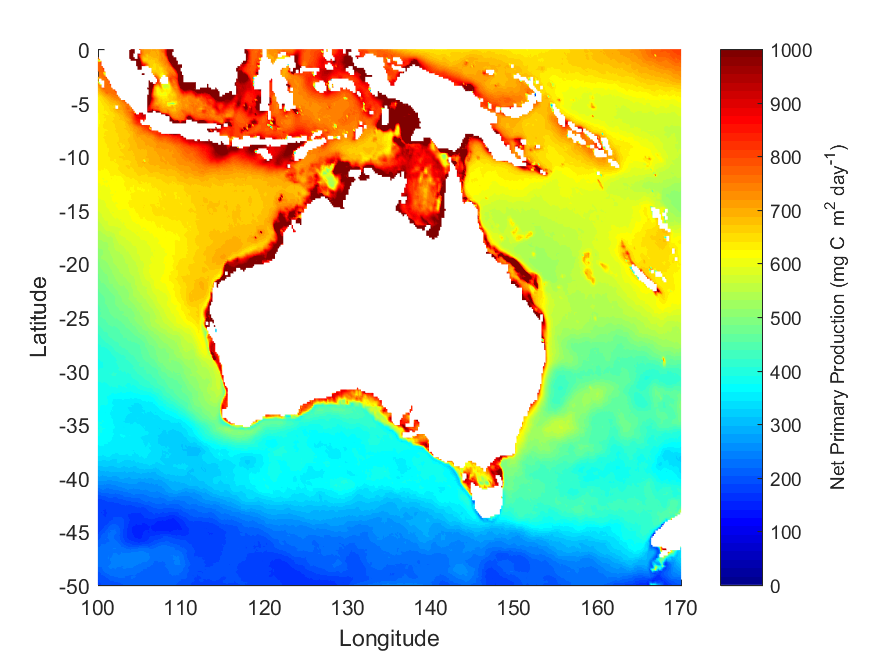
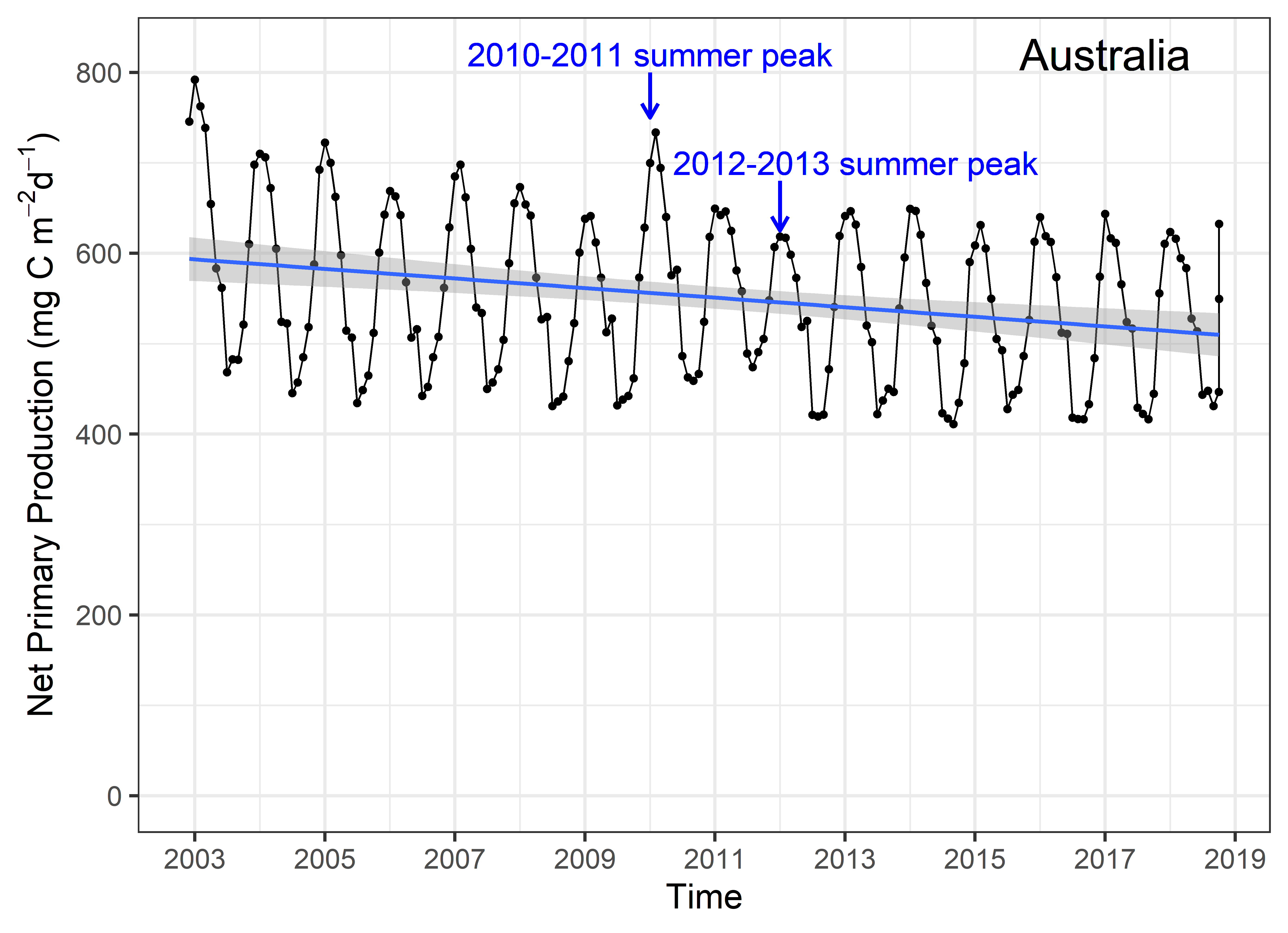
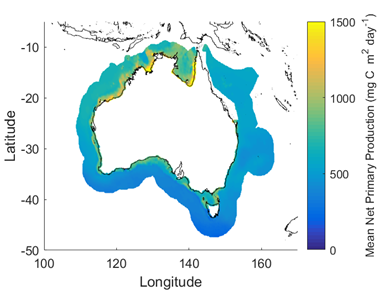
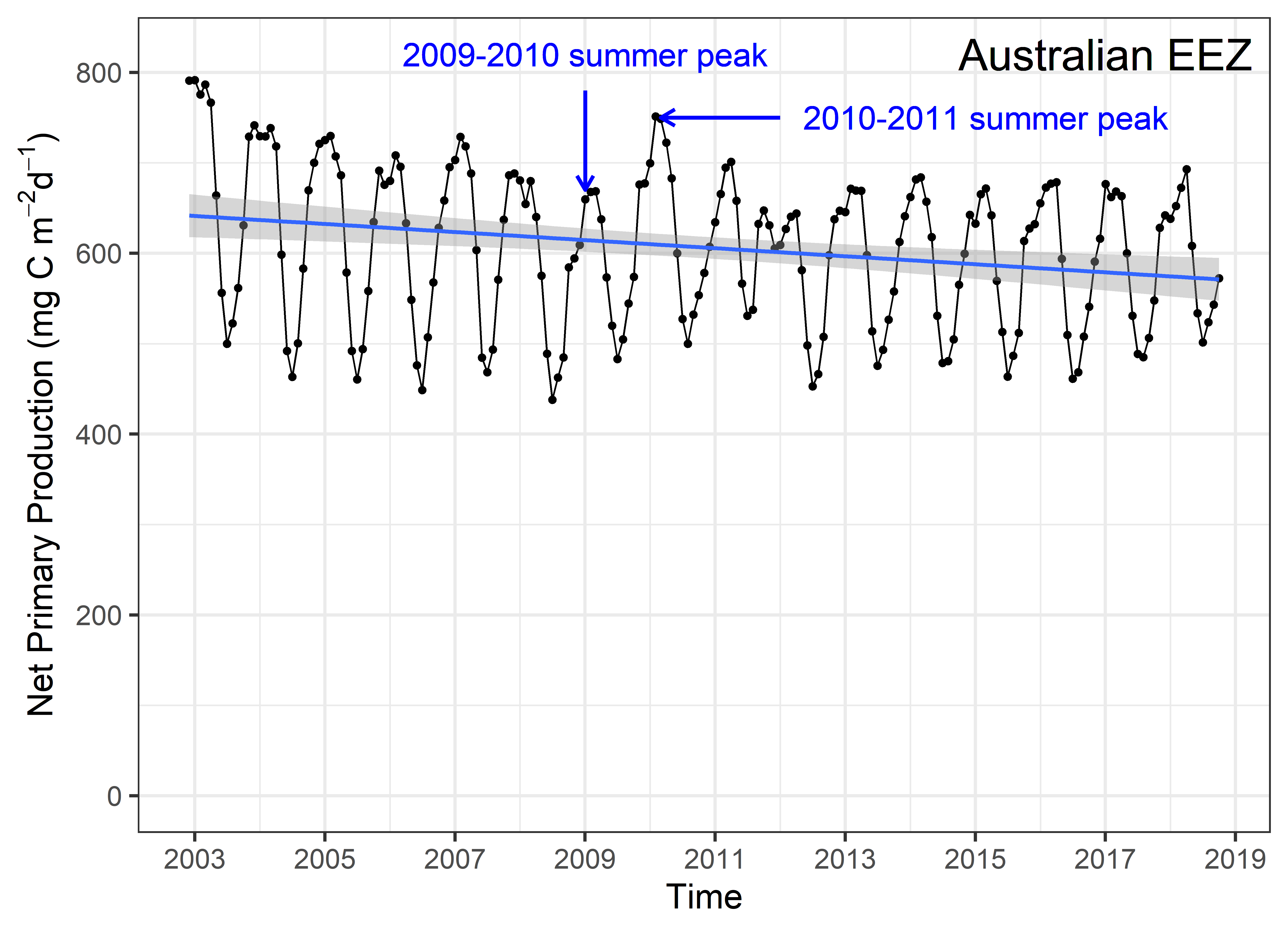
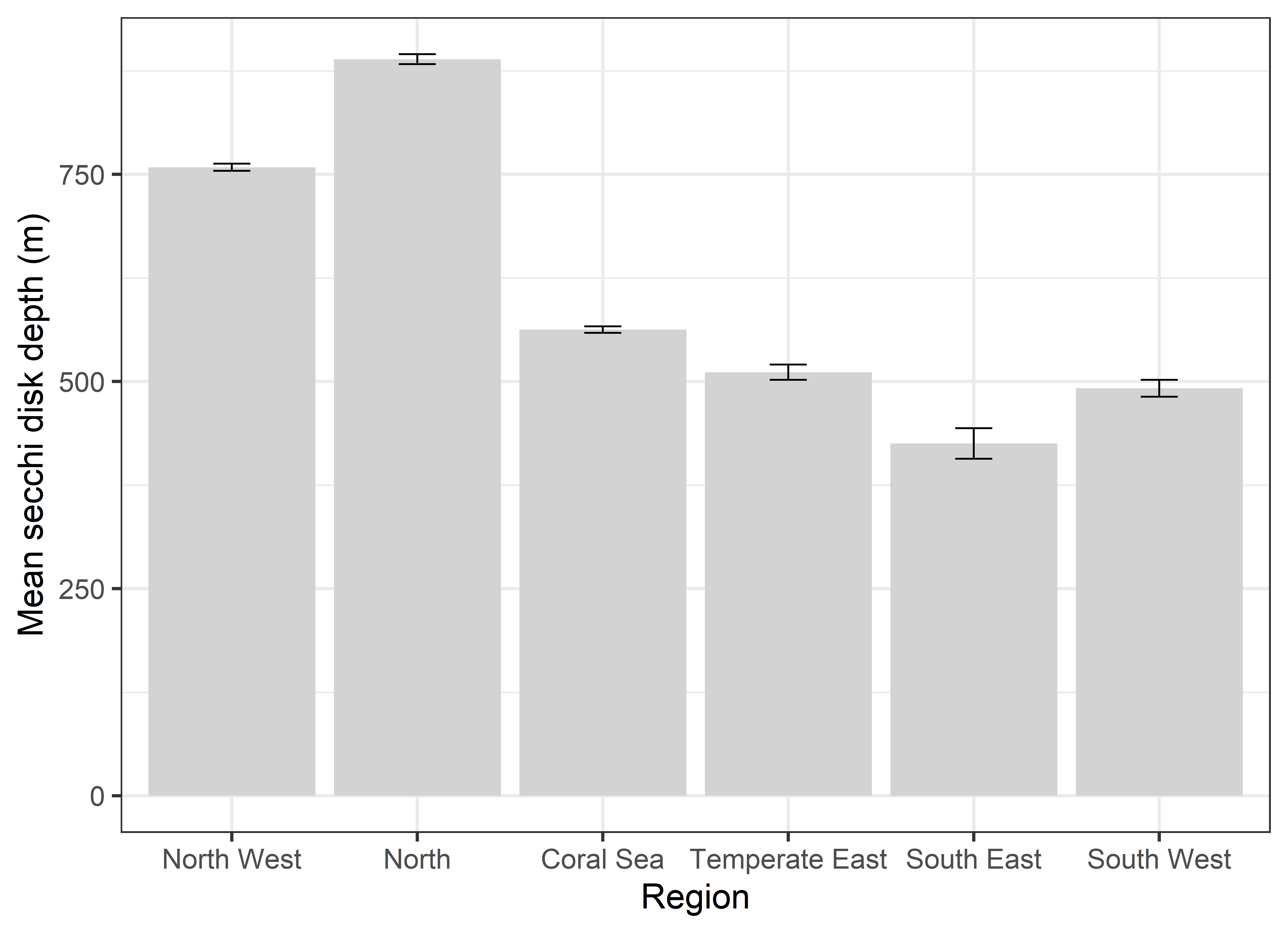
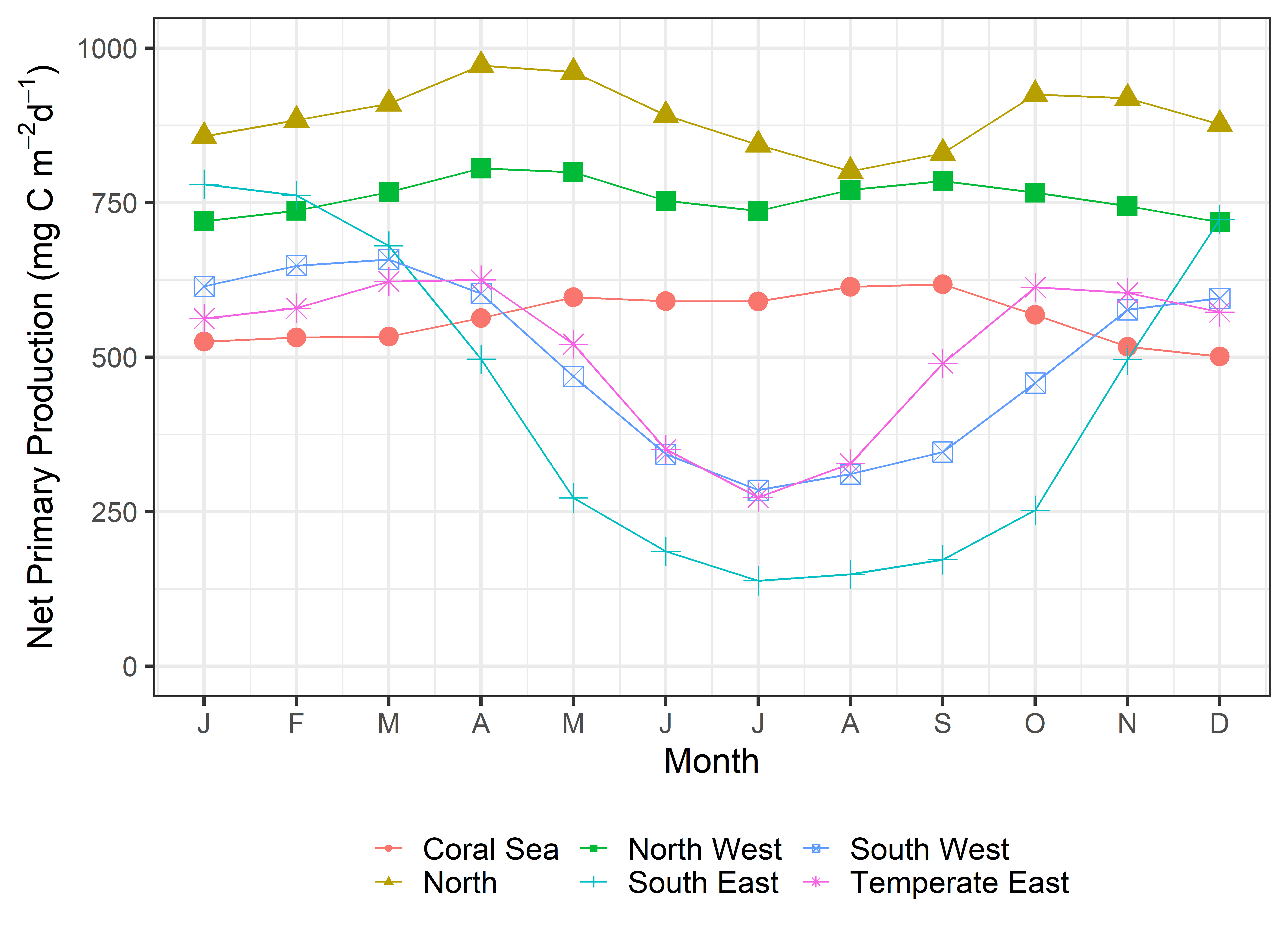
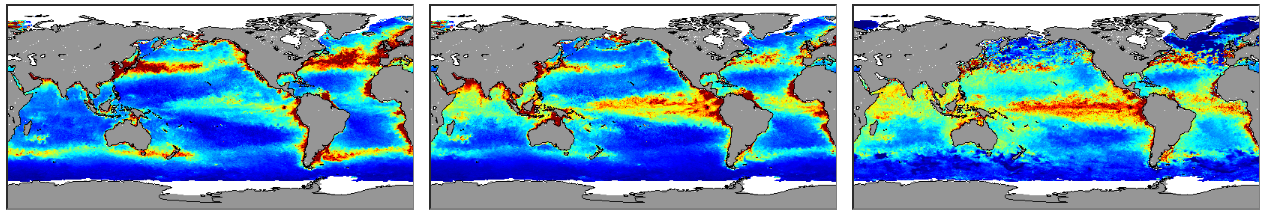
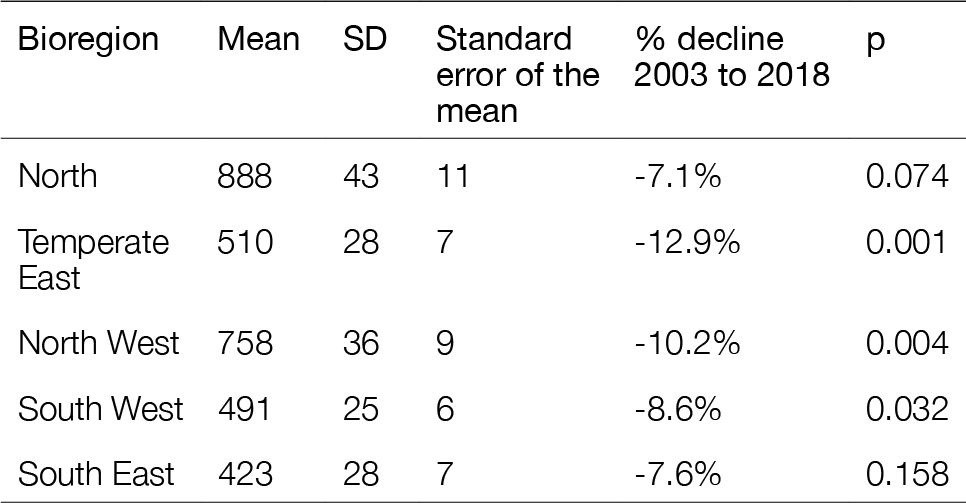
Net primary productivity varies greatly around Australia, with highest rates in coastal zones and in seas between Australia and its northern neighbours. Although there is a strong seasonal trend, increasing during spring-summer and decreasing in autumn-winter, average net primary productivity has declined in Australian waters by 12 % in the past 17 years (2002-2019). Given that primary production sets the carrying capacity of higher trophic levels (zooplankton, fish, birds and mammals), this decrease is likely to cause a reduction in the size of Australia’s fisheries.
Keywords
secchi disk depth, carbon-based, variability
Spatial and seasonal trends in net primary production
Download this Time Series Report
Citing this report:
Thompson P, McDonald K. (2020). Spatial and seasonal trends in net primary production. In Richardson A.J, Eriksen R, Moltmann T, Hodgson-Johnston I, Wallis J.R. (Eds). State and Trends of Australia’s Ocean Report. doi: 10.26198/5e16a4a749e7a
doi: 10.26198/5e16a4a749e7a


Citing the Report
Richardson A.J, Eriksen R, Moltmann T, Hodgson-Johnston I, Wallis J.R. (2020). State and Trends of Australia’s Ocean Report, Integrated Marine Observing System (IMOS).
The State and Trends of Australia's Ocean Report was supported by IMOS. IMOS gratefully acknowledges the additional support provided by the Commonwealth Scientific and Industrial Research Organisation (CSIRO).
The State and Trends of Australia's Ocean website is maintained by IMOS.

Australia’s Integrated Marine Observing System (IMOS) is enabled by the National Collaborative Research Infrastructure Strategy (NCRIS). It is operated by a consortium of institutions as an unincorporated joint venture, with the University of Tasmania as Lead Agent.
Disclaimer:
You accept all risks and responsibility for losses, damages, costs and other consequences resulting directly or indirectly from using this site and any information or material available from it. While the Integrated Marine Observing System (IMOS) has taken reasonable steps to ensure that the information on this website and related publication is correct, it provides no warranty or guarantee that information provided by the authors is accurate, complete or up-to-date. IMOS does not accept any responsibility or liability for any actions taken as a result of, or in reliance on, information on its website or publication. Users should check with the originating authors to confirm the accuracy of the information before taking any action in reliance on that information.
If you believe any information on this website or in the related publication is inaccurate, out of date or misleading, please bring it to our attention by contacting the authors directly or emailing us at IMOS@imos.org.au
Images and Information:
All information on this website remains the property of those who authored it. All images on this website are licensed through Adobe Stock, Shutterstock, or have permission from the original owner.